細胞分裂と細胞周期
特別な細胞を除いて、細胞はその内容物を倍加させて分裂して増える性質を持っています。酵母のような単細胞生物であるならば、細胞分裂自体が新しい個体を生み出す活動となります。
細胞が増殖する際にはある決められたサイクルがあり、それを細胞周期と呼びます。細胞周期では、DNAを複製を準備する期間(G1期)、DNAを複製する期間(S期)、細胞分裂準備期間(G2期)、細胞分裂期間(M期)の4つのステージがあります。哺乳類の細胞はだいたい1時間ほどでM期を終えることができます。
細胞周期はやみくもに進んでいるわけであなく、細胞周期制御系がそのサイクルを支配しています。制御系の中心となっているのは調節たんぱく質です。調節たんぱく質は遺伝子発現を促し、細胞周期を進めていきます。サイクリン依存タンパクキナーゼ(Cdk)はS期やM期への移行を制御しています。また、M-Cdk(M期Cdk)と呼ばれるキナーゼは、形態変化(染色体の凝縮、核膜の分散、小胞体とゴルジ体の作り直し、細胞外マトリックス接着の弛緩…)を引き起こし、細胞分裂になくてはならないトリガーとなる調節たんぱく質です。
染色体をまとめる:コヒーシンとコンデンシンの働き
有糸分裂を行う細胞にとって重要なことは、複製されたDNAを正確に2つに分離させることです。真核生物においては僅かな例外を除いて共通したシステムを持っており、細胞骨格を組み立てて装置を作り、染色体を引っ張ることで2等分します。M期に入る前には、染色体を平等に分配するために染色体を正しい形(通常はバラバラのひも状)に整えられます。それはS期に始まります。
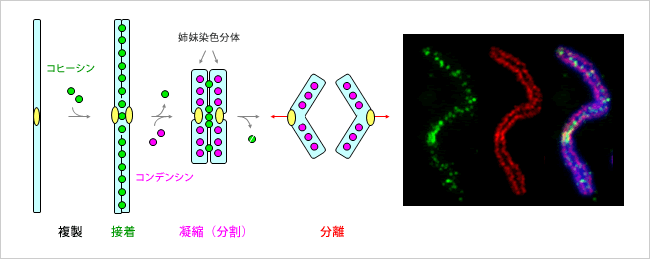
S期では複製された染色体は姉妹染色分体として固く結合しています。コヒーシンと呼ばれるたんぱく質が、姉妹染色分体の全長にわたって、染色体同士を結合させています。また、コンデンシンと呼ばれるたんぱく質が働き、染色体凝縮を行います。つまり、一本のひも状となっているDNAを巻き上げ、染色体を小さくコンパクトな形状にします。
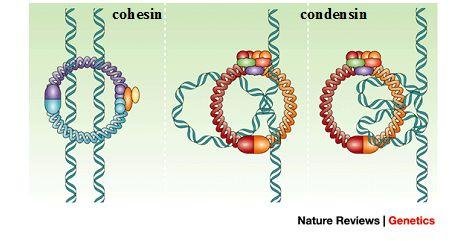
コヒーシンもコンデンシンも構造が似ているので混乱しやすいですね。コヒーシンは姉妹染色分体をひとくくりにする、コンデンシンは姉妹染色分体それぞれにおいてぎゅーっと凝縮させる役割、と理解すると良いでしょう。下画像が示す通り、まず初めにコピー同士を結合させ、それからコヒーシンが外れながら、左右に姉妹染色分体をコンデンシンによってコンパクトにしていきます。コヒーシンが全て外れるのは分裂するときです。
http://www.riken.jp/chromdyna/
中心体が2つに分かれる
複製された染色体が凝縮すると、細胞骨格が両極から次々に形成されていきます。まるで糸が伸びてくるようなので、有糸分裂と呼びます。中心体から細胞骨格が伸びた装置を有糸分裂紡錘体と呼び、DNAを正確に二分するという大切な役割を持っています。
また、染色体が分かれた後には細胞質も分離します。動物細胞においては、この分離は収縮環と呼ばれるアクチンフィラメントの輪っかが細胞質をしぼるようにしてちぎって分裂させます。植物細胞では細胞壁があるため、細胞板を間に形成するという、全く異なる分離方法をとっています。
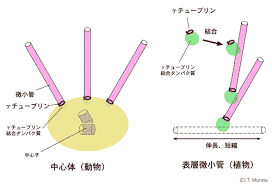
M期が始まる前には中心体が2つに増加します。中心体は、微小管形成中心であり、有糸分裂紡錘体の2つの極(基地のようなもの)を形成します。中心体はたくさんのγ-チューブリン環を持っており、放射状に伸びる微小管の伸長の基点となります。
動物の細胞周期の間期にはすでに中心体は2つに倍加していますが、分離はしていません。M期に入ると分離が開始され、微小管が放射状に並んだ構造となります。これを星状体と言います。星状体は、両側に移動し、染色体の中心体に結合して、紡錘体を形成します。DNA複製の引き金となるのと同じCdkが中心体の引き起こすことも知られており、S期の初期には中心体の複製が始まっていると考えてよいでしょう。
有糸分裂紡錘体を形成する
紡錘体によって左右両側からひっぱられた染色体は、2等分されます。この2等分された染色体を娘染色体と呼びます。
微小管はチューブリンを単位とする付加と解離によってたえず重合と脱重合を繰り返している状態にあり、これを動的不安定と呼びます。有糸紡錘体が形成される際には、細胞質中の微小管が分解(脱重合)し、一方で有糸紡錘体の微小管に集合(重合)します。分裂の際には有糸紡錘体の微小管の脱重合が起こり、染色体が両極に引っ張られていきます。
この微小管の伸縮は微小管付随たんぱく質(MAP)の活性の変化によって生じることがわかっており、M期の引き金となるM-CdkがMAPをリン酸化させ、微小管の安定性を低下させ、細胞質中の微小管の脱重合が進みます。また、カタストロフィンと呼ばれるたんぱく質も大規模な脱重合をさらに進め、有糸紡錘体の微小管形成のための材料(チューブリン)が確保されます。
複製された2個の中心体はモータータンパク質によって両極に移動され、中心体からはあらゆる方向へ微小管が伸びだします(重合と脱重合を繰り返す)。そのうち、伸びた微小管の一部が反対側の微小管と接触するようになり、その相互作用によって安定し、重合が防がれる働きが生まれます。その結果、左右の中心体が結ばれ、安定的な有糸分裂紡錘体を形成することができるのです。ちなみに、この紡錘体に含まれる2個の中心体は紡錘体極、微小管を極間微小管と呼びます。
植物の場合では中心体はありませんが、紡錘体を形成することはわかっています。染色体が核となって微小管を集合させ、モーターたんぱく質が微小管と染色体をつなぎ合わせながら紡錘体を作っていきます。
有糸分裂紡錘体が染色体と結合する
始めに、核膜が消滅する反応が起こります。これが引き金となるのは核ラミナに存在する中間径フィラメントのリン酸化と、分散です。核ラミナは核膜の内側にある細胞骨格の網目構造で、核膜の構造を支持する働きを持っています。これが、リン酸化によて分散し、核膜は小さい膜小胞となります。
その結果、染色体が現れ、微小管が染色体の動原体と呼ばれるたんぱく質複合体に結合します。このような動原体に結合する微小管は、極間微小管と区別して動原体微小管と呼びます。動原体は、染色体の中心にある特徴的なくびれ(セントロメア領域)に存在しており、セントロメア領域にはある特徴的な塩基配列が組み込まれています。動原体は姉妹染色分体どちらにもあるため2つあります。
動原体に付着する微小管の数は生物種によって異なり、ヒトの場合には1つの動原体に20-40本の微小管が結合することが知られています一方、酵母は動原体に1本の微小管しか結合しません。