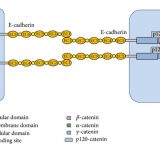
なぜ二重膜なのか
原核生物同士が共生して、ミトコンドリアや葉緑体になったという説を共生説(細胞内共生説)という。ミトコンドリアや葉緑体を真核生物の膜が包みこむようにして共生を果たしたと考えられており、それが二重膜の起源とされている。

ミトコンドリアの異質二重膜
共生説を支持する根拠として、ミトコンドリアの内膜と外膜の成分が違うことが挙げられており、これを異質二重膜と呼ぶ。

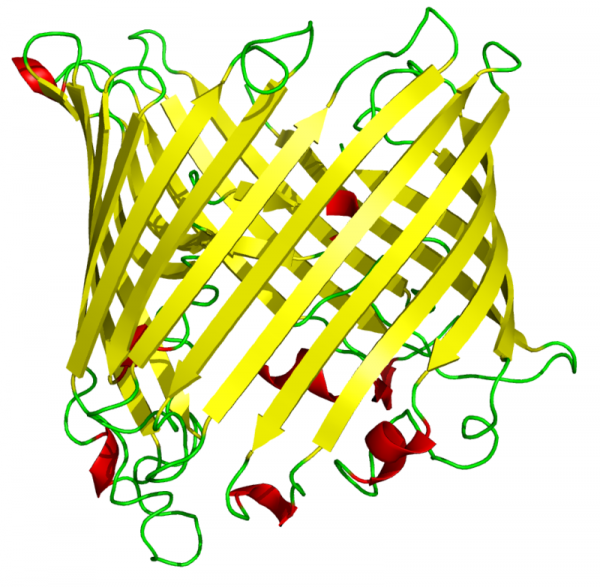
外膜にはポリンと呼ばれる膜タンパク質が大量にあり、分子量5000以下の分子が自由に行き来できるチャネルを形成している。一方、内膜は外膜とは対照的に不透過性で、物質を輸送するために輸送体が必要となる。マトリックスへのタンパク質輸送装置や、ミトコンドリア分裂・融合に関するタンパク質などが存在し、ミトコンドリアの全タンパク質の2割が含まれる。
膜成分の重量比としては次のような違いがある。
| ミトコンドリア外膜 | タンパク質:リン脂質=1:1(細胞膜と同じ) |
| ミトコンドリア内膜 | タンパク質:リン脂質=3:1 |
葉緑体はシアノバクテリア由来の二重膜(かもしれない)
教科書においては、ミトコンドリア外膜・葉緑体外膜どちらも真核生物(寄生先)由来との扱いであるが、最近の研究によると葉緑体外膜・内膜ともにシアノバクテリア由来の同質膜であるらしい。
葉緑体の二重包膜は、現在はいずれも共生したシアノバクテリアの二重膜を起源とすると考えられています。真核生物がシアノバクテリアを細胞内共生で取り込んだ際、取り込まれたシアノバクテリアは宿主の食包膜に囲まれていたと想定されますが、共生の初期段階でこの食包膜は消失したと考えられています。
最近の研究から、葉緑体外包膜とシアノバクテリアの外膜は、いくつかの点で共通していることが判ってきました。例えば、外膜を構成するガラクト脂質や葉緑体へのタンパク質輸送に働く外包膜の輸送体(TOC複合体の一つであるTOC75 βバレルタンパク質)がどちらの膜にも共通して存在します。最近の分子系統解析から単系統であるということが示唆されている灰色植物の葉緑体も2重膜であること(これはその2枚の間にペプチドグリカンを持っていますのでシアノバクテリアの膜構造と基本的に一致します)も外膜がシアノバクテリア由来であることを示唆します。
しかし、このような知識は未だ一般化しているとは言えないため、当分の間はミトコンドリア外膜も葉緑体外膜も真核生物由来の膜であると覚えておいて良い。