ねこでもわかる生物学
生物学Q&A
生物学まとめ
生物本
やばい生き物
生き物雑学
ABOUT
ちょっとディープな生物の世界
ホーム
「生命現象とタンパク質」の記事一覧
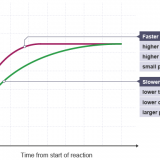
酵素に関する生成物量・反応速度のグラフ-競争的阻害・非競争的阻害・アロステリック効果の違い-
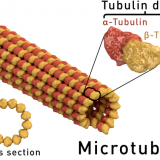
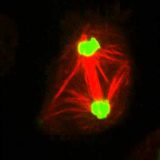
微小管による運動-鞭毛の構造と曲がる仕組み-
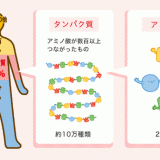
生体内のタンパク質の種類
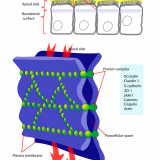
細胞接着:密着結合、接着結合、ギャップ結合、デスモソームによる結合の違いとは?
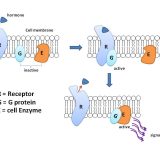
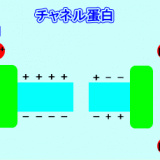
内分泌に関するタンパク質-Gタンパク質・Gタンパク質共役型受容体・ステロイドホルモン受容体-
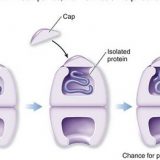
細胞内のタンパク質-シャペロン-
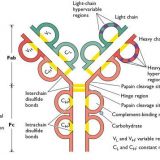
細胞内のタンパク質-免疫に関するタンパク質-
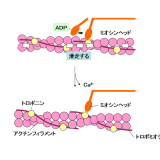
細胞内のタンパク質-モータータンパク質-
細胞内のタンパク質-神経細胞-
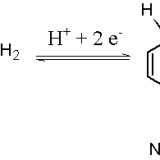
補酵素と反応の調節
酵素の反応:基質特異性・最適温度・最適pH・競争的阻害・非競争的阻害
タンパク質の構造とアミノ酸の種類